Bernhard
Peter
Der
Vermehrungszyklus von HIV
Vermehrungszyklus
von HI-Viren
Die Vermehrung
von HI-Viren kann
man in mehrere Schritte unterteilen:
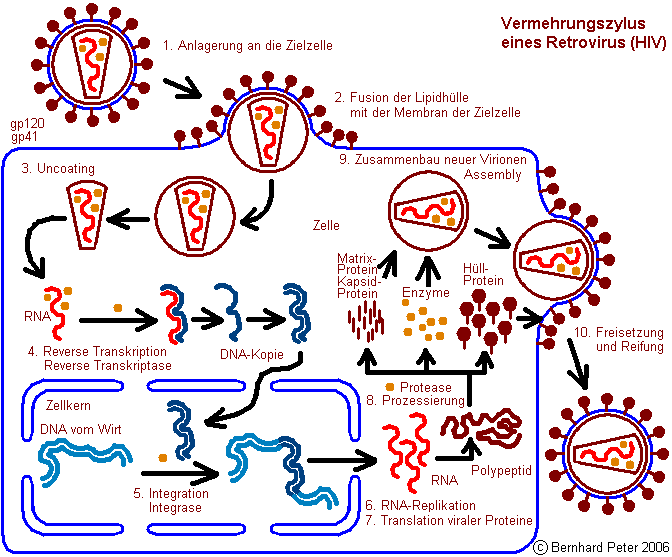
1. Anlagerung an die Zielzelle
2. Fusion der Lipidhülle mit der Membran der Zielzelle
3. Uncoating
4. Reverse Transkription
5. Integration
6. RNA-Replikation
7. Translation viraler Proteine
8. Prozessierung
9. Assembly – Zusammenbau neuer Virionen
10. Freisetzung und Reifung
Schritt
1:
Anlagerung an die Zielzelle
Entscheidend für
den Erfolg einer
Infektion ist das „Matching“ zwischen Virus und
Zielzelle. In seine Lipidhülle eingebaut trägt das
Virus
Glykoproteine, die ihm spezifische Oberflächeneigenschaften
verleiht. Erst wenn es zu einer Wechselwirkung mit passenden
Oberflächenstrukturen einer Zelle kommt, kann sich der Virus
anlagern und im nächsten Schritt aufgenommen werden.
Im Detail müssen zusammenpassen: Seitens des Virions das Glykopeptid gp120 sowie seitens der Zelle CD4-Peptid (zuerst) und CCR5-Chemokin-Rezeptor (danach). Dieses Prinzip erklärt, warum die Viren nur ganz bestimmte Zielzellen befallen, im Falle des HIV eine bestimmte Sorte der T-Helferzellen, nämlich die mit dem Eiweiß namens CD4, sowie die Monozyten mit den selben Oberflächen-Strukturen.
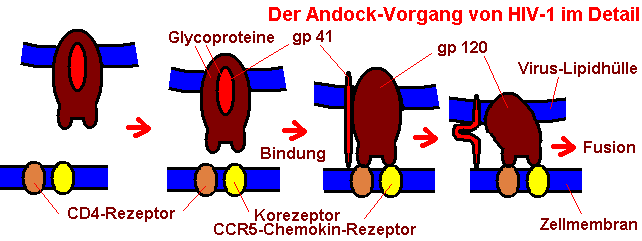
CD4 befindet sich auf der Oberfläche von ca. 60% aller T-Lymphozyten, von T-Vorläuferzellen in den immunogenen Organen, dazu auf Makrophagen, Monozyten, eosinophilen Leukozyten, Mikrogliazellen, dendritischen Zellen etc. Die erste Kontaktaufnahme zwischen Virion und menschlicher Zelle erfolgt über eine Wechselwirkung von dem CD4-Protein mit dem gp120-Glykoprotein. Doch dieser Prozeß leitet die Bindung erst ein, denn für eine vollständige Bindung ist eine zweite Wechselwirkung notwendig, nämlich die des gp120-Glycoproteins mit einem Korezeptor, in den meisten Fällen ist es der CCR5-Chemokin-Rezeptor für die M-tropen Viren (monozytotropen Viren), dagegen der CXCR4-Korezeptor bei T-tropen Viren (T-zell-tropen Viren).
Schritt
2:
Fusion der Lipidhülle mit der Membran der Zielzelle
Retroviren sind
behüllte Viren –
eine Lipiddoppelmembran umgibt die Kapside wie ein Ballon oder
wie eine Seifenblase. Das Grundmaterial für die
Lipiddoppelmembran, die die Viren umgibt, ist genau das gleiche
wie das der Wirtszelle, einmal von den spezifischen, in die
Membran eingebauten Glycoproteinen abgesehen. So wie zwei
Seifenblasen miteinander zu einer größeren
fusionieren können,
so fusionieren auch Viren-Lipid-Membran und
Wirtszell-Lipid-Membran.
Im Detail ist das ein wenig komplexer: Wenn das virale Glycoprotein gp120 an die zelleigenen Transmembranproteine CD4 und CCR5 gebunden hat, dann kommt es zu einer Konformationsänderung der Glycoproteine. Das gp120 weicht zur Seite, das bisher darunter verborgene gp41 kommt hervor und verhakt sich mit seinem N-terminalen Ende in der Zelloberfläche. Dann sorgt das virale Glycoprotein gp41 dafür, daß sich Virus und Zelle so nahe kommen, daß die Fusion stattfindet, indem zwei Abschnitte von gp41 freigelegt werden, die sich miteinander verbinden. Erst infolge dieser Konformationsänderung kann die virale Membran mit der der Zielzelle fusionieren. Im Ergebnis findet sich das Viruskapsid mit dem genetischen Material im Zellinnern wieder, aber ohne Hülle.
Schritt
3:
Uncoating
In diesem Schritt wird das
Virion
weiter zerlegt, das Kapsid wird abgetrennt, letztendlich bleibt
nur die Information und die Funktion übrig: Die Virus-RNA und
die Virus-Enzyme.
Schritt
4:
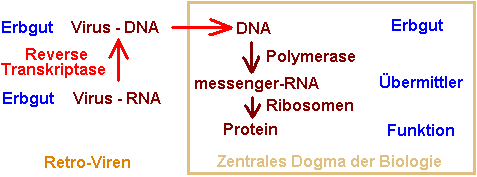
Reverse Transkription - Retroviren kehren das zentrale Dogma der
Biologie um
Bis zum Bekanntwerden war
für
Molekularbiologen die Welt noch in Ordnung: Man hatte ein
„zentrales Dogma“, welches besagte, daß
der
Informationsfluß in der Zelle ganz klar in der Hierarchie
gerichtet ist: Die DNA ist die Erbinformation, deren Information
von RNA übermittelt wird, damit Proteine (Eiweiße)
in der Zelle
synthetisiert werden. Der Informationsfluß erfolgt also von
der
Leitzentrale über die Boten zu den Ausführenden, von
der DNA
über die RNA zu den Eiweißen.
Und das war mit einem Mal nicht mehr gültig! Die Retroviren durchbrachen dieses sog. zentrale Dogma der Molekularbiologie: Der Virus schleust RNA ein – wie viele andere Viren auch, aber diese anderen herkömmlichen RNA-Viren beschränken sich darauf, über ihre RNA den Eiweißstoffwechsel der Zelle in ihrem Sinne zu verändern. Retroviren dagegen verfügen über ein spezielles Enzym, welches das bisher für unmöglich Gehaltene wahr macht: Die sog. Reverse Transkriptase schreibt die Einzelstrang-RNA des Virus in eine DNA um! Damit ist die Richtung des Informationsflusses umgekehrt worden.
Im darauffolgenden Schritt agiert die Reverse Transkriptase wie eine Nuklease: Der RNA-Strang wird entfernt, und zur der bislang einzelsträngigen neuen DNA wird ein komplementärer Strang synthetisiert, so daß man doppelsträngige provirale DNA erhält.
Exkurs:
Endogene Retroviren:
Retroviren sind sogar ein
ganz normaler
und meist unauffälliger Bestandteil des menschlichen Genoms.
Sie
sind als gesammelte Spuren früherer Infektionen, die
während
der Evolution Zellen der Keimbahn betrafen, übriggeblieben,
und
sie werden als „Datenmüll“ seitdem in der
Regel
folgenlos weitervererbt – insgesamt ca. 8 % (!) des
menschlichen Erbgutes, 26 verschiedenen Linien zuzuordnen, die
jeweils auf einzelne Infektionen zurückgehen. Die Erinnerung
daran wird die Spezies Mensch nie mehr los.
Schritt
5:
Integration
Ein weiteres Enzym,
welches der Virus
mitbringt, ist die Integrase. Die sorgt dafür, daß
die neue DNA
nach Transport in den Zellkern in die DNA-Erbinformation der
entsprechenden Chromosomen der Wirtszelle eingebaut wird. Der
Einbau ist notwendig, damit die Virus-DNA wie bei der normalen
DNA abgelesen und Messenger-RNA hergestellt werden kann. Zu
beiden Seiten des Virus-Gens befindet sich eine sog LTR-Zone
(long terminal repeat) – das sind die Schnittstellen, die
beidseitig mit der DNA des Wirtes verbunden werden. Erst die
Aktivierung der CD4-Zelle ermöglicht die Integration. Das
eigentlich Fatale an dem Einbau ist: Die Wirtszelle kann sich nie
mehr von der Virus-DNA trennen, weil diese fester Bestandteil der
eigenen Erbinformation geworden ist! Schlimmer noch: Die Zelle
kann auch nicht erkennen, welche Abschnitte eigene und welche
fremde Informationen enthalten! Der Zellstoffwechsel kann jetzt
vom Virus fremdgesteuert werden.
lebenslange
Reservoire
Solange die CD4-Zelle aber
nicht
aktiviert ist, erfolgt keine Integration. Das HIV-Genom liegt als
provirale, nicht-integrierte Virus-DNA vor. Latent infizierte und
ruhende CD4-Zellen mit nicht integrierter DNA stellen ein
lebenslanges Reservoir dar und machen die medikamentöse
Bekämpfung des Virus zu einer nie endenden Sisyphusarbeit.
Auch
Makrophagen, Monozyten und Mikrogliazellen etc. stellen ein
lebenslanges Reservoir dar, das z. T. unerreichbar ist.
Schritt
6:
RNA-Replikation
Wie vom normalen
Zellstoffwechsel
gewohnt, wird die DNA abgelesen, es werden RNA-Moleküle
gebaut.
Darunter ist vor allem RNA, die identisch ist mit derjenigen, die
die Zelle einst infiziert hat – die genetische Information
des HI-Virus. Es werden größere Mengen davon als
„Rohstoff“ für die Virusproduktion
hergestellt, dabei
wird auf die RNA-Polymerase der Wirtszelle zurückgegriffen,
eine
solche hat das Virion nicht mitgebracht.
Schritt
7:
Translation viraler Proteine
Willig folgt die Zelle den
auf dem
neuen Abschnitt kodierten Befehlen, welche Eiweiße zu bauen
sind
– Virus-Eiweiße natürlich. Für
neue Viren werden viele
verschiedene Eiweiße benötigt. Zum einen sind das
die
Kapsid-Proteine, aus denen das Kapsid besteht, dann die
Matrix-Proteine, mit denen die Hülle innen ausgekleidet ist,
dann die Proteine, die später als Glycoproteine in der
Lipidmembran der Hülle (envelope) eingebaut werden. Das waren
jetzt 3 Sorten Struktur-Proteine. Es werden aber auch 3
Funktionsproteine gebraucht, die die neuen Viren mit auf ihre
Reise zu neuen Zellen bekommen: Die Reverse Transkriptase, die
Integrase und die Protease.
Schritt
8:
Prozessierung
Die Protease wird
gebraucht, um das
Primärprodukt der Translation, die Vorläufer-Proteine
in die
einzelnen funktionsfähigen Stücke zu zerschneiden,
denn die
einzelnen Eiweiße werden erst einmal kontinuierlich gebaut
und
dann in die benötigten Einzelteile zerschnitten. Weitere
Schritte sind notwendig, um aktive Bauteile für die neuen
Viren
zu erhalten, das nennt man Prozessierung der Primärprodukte.
Schritt
9:
Assembly – Zusammenbau neuer Virionen
Viren sind meist so
gebaut, daß ihr
Kapsid aus wenigen repetitiven Einheiten besteht, die durch
Selbstassoziation bestimmte Raumelemente wie in Modul-Bauweise
bilden. Die hohe Regelmäßigkeit im Aufbau macht die
Assoziation
der einzelnen Elemente zu einem neuen Virus einfach.
Schritt
10:
Freisetzung und Reifung
Retroviren sind
behüllte Viren. Das
Material für die Lipiddoppelmembran, die die Viren umgibt,
stammt aus der Zellmembran der Wirtszelle. Bei der Freisetzung
wird ein Abschnitt der Zellmembran ausgestülpt und
abgeschnürt,
so daß er wie ein Ballon das Viruskapsid umgibt. Vorher
werden
aber in die Lipidmembran bestimmte Glycoproteine eingebaut, die
dem neuen Virus seine speziellen Oberflächeneigenschaften
verleihen. Diese Glycoproteine sind in der Membran der Wirtszelle
nicht vorhanden und werden in den Schritten 7 und 8 hergestellt.
Erst durch diese Glycoproteine erhält der neue Virus seine
spezifischen Oberflächenstrukturen, mit denen er neue Zellen
erkennen und infizieren kann. Die Viren verlassen die Wirtszelle
und infizieren weitere T-Helferzellen. Pro Tag können bis zu
10
Milliarden neuer Viren produziert werden – alles mit dem
Stoffwechsel des Patienten.
Je nach Zelltyp ist der Prozeß des „Budding“ unterschiedlich: Bei den T-Zellen werden die Viren gleich nach außen abgegeben, sie sprossen alle auf der Zelloberfläche. Aber bei Monozyten und Makrophagen ist es genau so möglich, daß sie erst in Membransysteme des Zellplasmas (Endoplasmatisches Retikulum, Vesikel) hinein gebildet und angesammelt werden und schließlich als ganze Gruppe an den Extrazellularraum abgegeben werden.
Literatur:
www.hiv.net
Fritz H. Kayser et. al., Medizinische Mikrobiologie, 11. Auflage,
Thieme Verlag ISBN 3-13-444811-4, S. 538-550
Zurück
zur Übersicht HIV / AIDS
Andere pharmazeutische
Seiten
![]() Home
Home
![]()
©
Copyright Text, Graphik und Photos: Bernhard Peter 2006
Impressum